コネクシンまたはコネキシン (Connexin) は脊椎動物に見られる一群の膜貫通タンパク質で、ギャップ結合を形成する。無脊椎動物のギャップ結合を形成するイネキシンとは構造的に関連がない。ギャップ結合は、コネクソンと呼ばれるヘミチャネル(半チャネル)の対から構成され、ヘミチャネルは6分子のコネクシンから構成される。心筋の協調した脱分極や胚の適切な発達、微小血管系での伝導反応等、多くの生理過程に必須であり、コネクシンをコードする遺伝子の突然変異は、機能や発達の異常に繋がる。
構造
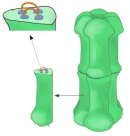
コネクシンは、C末端とN末端がともに細胞質側にある4回貫通型の膜貫通タンパク質で、細胞質ループ(CL)と2つの細胞外ループ(EL-1及びEL-2)からなる。コネクシン遺伝子ファミリーは多様であり、ヒトゲノムには21個、マウスのゲノムには20個の配列が同定されており、そのうち19個はオーソログ遺伝子である。これらの分子量は26-60 kDaで、平均で380アミノ酸長である。様々な組合せのコネクシンがギャップ結合を形成し、伝導性、サイズや電荷に対する選択性、開口電位・開口物質等の性質が少しずつ異なるものとなる。
命名
コネクシンはCxまたはCXと略称される。最近の論文では一般にその分子量によって命名され、例えば分子量26 kDaのものはCx26と呼ばれる。だが、生物種が異なるとオーソログタンパクの分子量が異なることがあり、例えばヒトのCx36は、ゼブラフィッシュのCx35と相同である。これはコネクシンを比較する際の混乱の元となることから、GJ (Gap Junction) 命名法と呼ばれる別の命名法が提案された。これはコネクシンをα型(GJA)とβ型(GJB)、さらにC、D、E型に分類し、これに同定番号を続けるものである。例えば、CX43/Cx43に対応するのは、GJA1/Gja1となる。2007年にヘルシンゲルで行われたギャップ結合会議における投票で、コネクシンをコードする遺伝子にはGJ命名法を用いることが決定されたが、ヒトタンパク質の分子量を用いて、そのオーソログとなるタンパク質を表すことで従来の命名法を残すことも希望された。
生合成と分解
コネクシンの際立った特徴の一つとして寿命が非常に短いということが挙げられ、半減期はわずか数時間しかない。これはギャップ結合が動的に置換され続けているということを意味し、子宮筋層等の場所における細かい生理学的調整を可能としていると考えられる。
核から細胞膜まで
リボソームにより翻訳されたコネクシンは小胞体の膜に挿入される。ここでフォールディングが行われ、2つの細胞外ループEL-1とEL-2が形成される。続いて分子がオリゴマー化し、ヘミチャネルが作られ始める。その後コネクシンは小胞体を出てVTC (Vesicular-tubular cluster) を経由し、シス・ゴルジ網に入る。だが、Cx26等、ゴルジ体を経由せずに輸送されるコネクシンもある。
ギャップ結合の組立て
細胞膜に挿入されたヘミチャネルは、脂質二重層中を自由に拡散する。主にカドヘリン等の特定のタンパク質の助けを借りて、ヘミチャネルは隣接細胞のヘミチャネルと結合し、ギャップ結合を形成する。近年の研究で、接着結合とギャップ結合の間の連絡が存在することが示され、より高度な調整が行われていることが示唆される。
機能
コネクシンは脊椎動物のみで見られる。無脊椎動物で同等の機能を果たすタンパクとしてイネキシンがあるが、これはコネクシンと遺伝的に関連がない。イネキシンのオーソログは脊索動物でも同定されているが、ギャップ結合は形成しない。これらのタンパク質で形成されたパネキシンと呼ばれるチャネルは、細胞の内外を繋ぐ非常に大きな膜貫通孔として働く。
中枢神経系内で、ギャップ結合は前駆細胞・神経細胞・グリア細胞間を電気的に共役する。特定のコネクシンを不活化したノックアウトマウスを用いた研究で、視覚信号の伝達にはこれらの電気的共役が必須であることが明らかとなった。網膜内では、周囲の光強度がギャップ結合による細胞共役に影響を与え、照明条件に応じて視覚機能を調節する。これらの電気的共役はコネクシンの発現も含め、様々な機構により制御される。
ヒトのコネクシンの一覧
出典
外部リンク
ウィキメディア・コモンズには、コネクシンに関するメディアがあります。
- Connexins - MeSH・アメリカ国立医学図書館・生命科学用語シソーラス(英語)